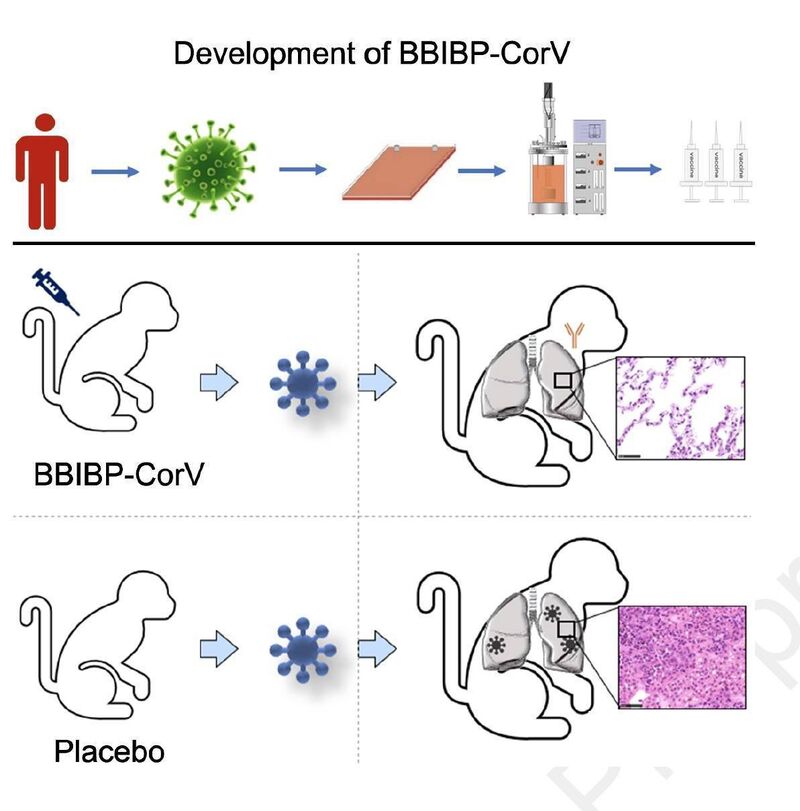
新冠大流行威胁着全球公共卫生。为防控新冠疫情,人类迫切需要疫苗。来自中国科研团队研发了名为BBIBP-CorV的新冠灭活疫苗,已在动物实验显示出了巨大的潜力:该疫苗高效、安全,在小鼠、大鼠、豚鼠、兔子和非人灵长类动物(食蟹猴猴子和恒河猴)体内均能诱导高水平的中和抗体滴度,2μg/剂便能提供针对新冠病毒的高效保护。同时,该候选疫苗是国内开发的第二个重要灭活疫苗,增加了国内的候选疫苗选择。
以上成果来自当地时间6月6日国际权威学术期刊《细胞》(CELL)在线发表的重磅论文:《开发候选灭活疫苗BBIBP-CorV,对SARS-CoV-2能提供有效保护》(Development of an inactivated vaccine candidate, BBIBP-CorV, with potent protection against SARS-CoV-2)。
该论文通讯作者为中科院院士、中疾控主任高福、中国医学科学院医学实验动物研究所所长秦川、中国疾控中心病毒病所应急技术中心主任谭文杰、清华大学教授娄智勇、中国食品药品检定研究院李长贵研究员、中国疾控中心生物安全首席专家、病毒病所党委书记武桂珍等。
研究团队来自北京生物制品研究所有限责任公司、中国疾病预防控制中心传染病预防控制所、国家人类疾病动物模型资源中心、国家卫生健康委员会人类疾病比较医学重点实验室、中国医学科学院医学实验动物研究所、北京协和医学院比较医学中心、新发再发传染病动物模型研究北京市重点实验室、清华大学等。
研究者详述了SARS-CoV-2灭活候选疫苗(BBIBP-CorV)的实验性生产,该疫苗在小鼠、大鼠、豚鼠、兔子和非人灵长类动物(食蟹猴猴子和恒河猴)体内均能诱导高水平的中和抗体滴度,以提供对SARS-CoV-2的保护。在恒河猴的气管内免疫接种中,研究使用2μg/剂量的BBIBP-CorV便可提供了对SARS-CoV-2的高效保护,同时没有检测到抗体依赖性增强(ADE)感染。此外,BBIBP-CorV在疫苗生产中表现出高效的生产力和良好的遗传稳定性。
与SARS-CoV和MERS-CoV相比,SARS-CoV-2似乎传播更快,导致对疫苗的迫切需求。迄今为止,三种候选疫苗(包括一种灭活疫苗、一种腺病毒载体疫苗和一种DNA疫苗)被报道以不同的效果保护恒河猴对抗SARS-CoV-2。灭活疫苗广泛应用于新发传染病的预防,其研制速度较快,针对新冠病毒的预防具有较好的开发前景。值得注意的是,新出现的证据显示,SARS-CoV感染的可能存在抗体依赖性增强(ADE),提示研制冠状病毒疫苗时应特别重视安全性评价。
国内的首个灭活新冠疫苗来自国药集团中国生物。国药集团中国生物4月14日发布消息,公司旗下新型冠状病毒灭活疫苗获得临床试验后,一期临床已在河南焦作地区开展。
另一方面,由军事科学院军事医学研究院生物工程研究所陈薇院士团队和中国康希诺(CanSino)公司联合开发的腺病毒5型(Ad5)载体疫苗也取得了重大进展。当地时间5月22日晚间,顶级医学期刊《柳叶刀》(The Lancet)刊发一篇论文,题为“重组腺病毒5型载体COVID-19疫苗的安全性,耐受性和免疫原性:剂量递增,开放标签,非随机,首次人类试验”。陈薇等研究人员报告了在中国健康成年人中使用的Ad5载体COVID-19疫苗在接种后28天内的1期临床数据,初步评估疫苗的安全性、耐受性和免疫原性。
疫苗设计和生产
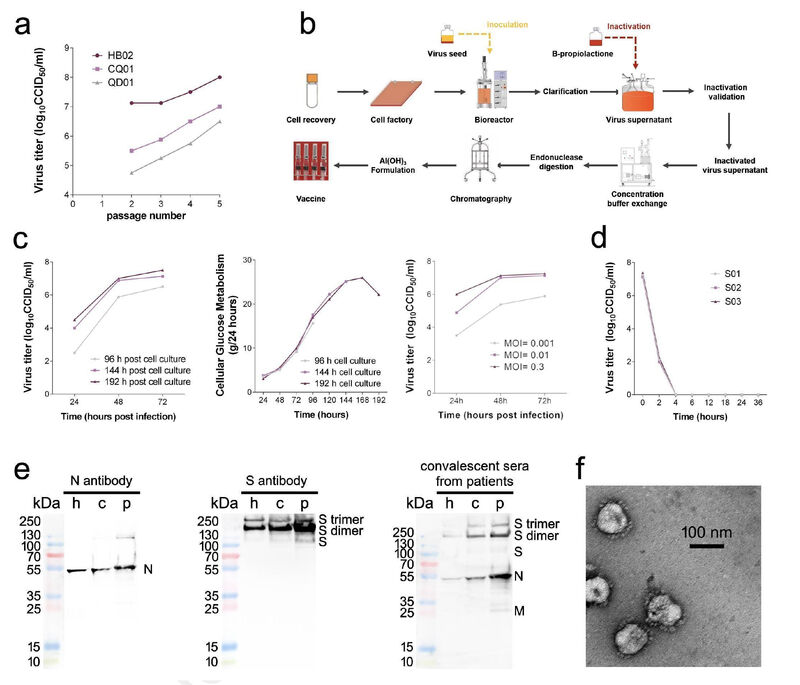
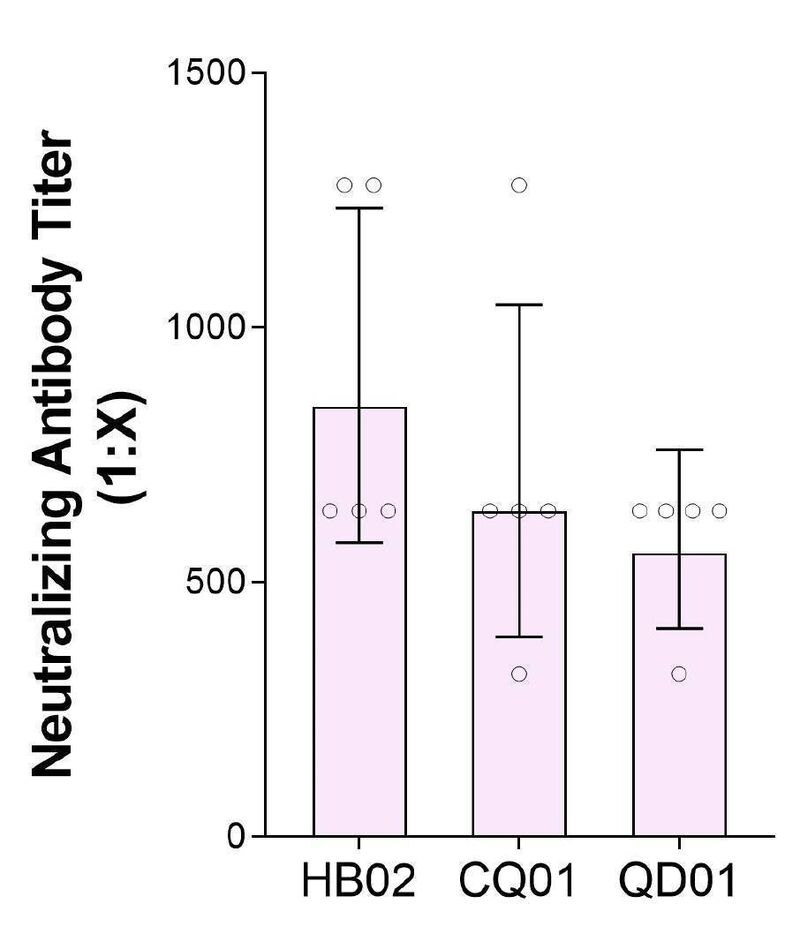
研究团队从3名新冠住院患者的支气管肺泡灌洗样本及咽喉拭子中分离出3株SARS-CoV-2毒株,用于建立SARS-CoV-2灭活候选疫苗的临床前体外中和攻毒动物模型。3株新冠毒株分别是:19 ncov-cdc-tan-hb02 (HB02), 19 ncov-cdc-tan-strain03 (CQ01)和19 ncov-cdc-tan-strain04 (QD01)。这3株毒株散布在系统发育树的不同位置,表明其对SARS-CoV-2具有较好的代表性。
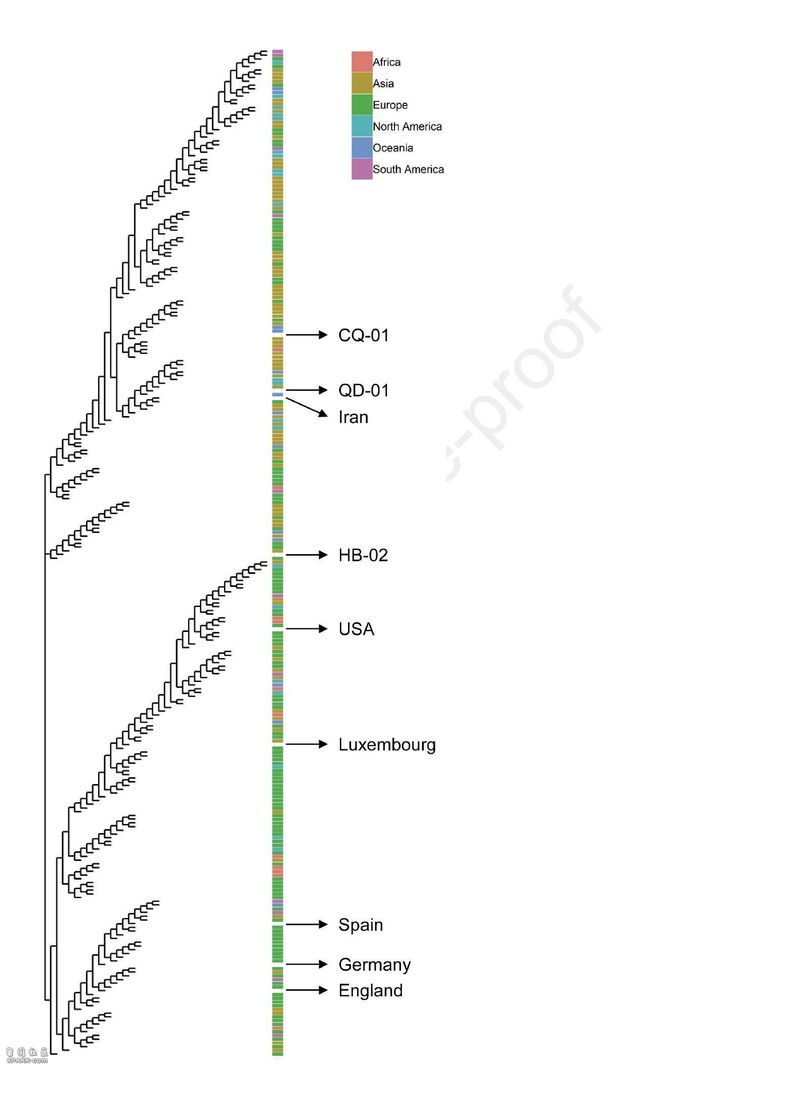
值得注意的是,所有这些毒株都是从Vero细胞中分离出来的,并已获得世界卫生组织的疫苗生产认证。这些Vero细胞通过患者的咽喉拭子感染,而不是其他细胞系,以防止在病毒培养和分离过程中可能发生的突变。高效增殖和高遗传稳定性是研制灭活疫苗的关键。他们首先发现在三种病毒株中,HB02株的88个Vero细胞复制效果最佳,产生的病毒产量最高(图1a)。因此,研究者选择HB02株进一步开发灭活SARS-CoV-2疫苗(BBIBP-CorV)。HB02株和其他从国内和国际来源的SARS CoV-2毒株的全基因组序列的比较表明:HB02毒株与其他病毒株同源,并证明了主要保护性抗原(刺突蛋白质)有100%的同源性。这说明具有广泛对抗各种SARS-CoV-2毒株的潜力(补充图1和补充图2)。为了获得适于高产的病毒培养基,研究者对HB02株进行了纯化,并在Vero细胞中传代产生了P1病毒培养基。将P1培养基在Vero细胞上进行适应性培养、传代和扩增。研究者将经过7代适应化的毒株(BJ-P-0207)作为生产疫苗的原种子(BJ-P1)。为评价遗传稳定性,研究者对其再传代3次,得到P10病毒培养基。通过深度测序分析,我们对HB02株和P10株进行了全基因组测序,结果显示它们的序列同源性大99.95%。此外,在P10原料病毒的全序列中,包括弗林分裂位点附近也没有发现氨基酸变异,这些结果表明HB02毒株具有较高的遗传稳定性,有利于进一步的发展。
为了高效制造,研究者建立了基于新型篮式反应器载体的BBIBP-CorV的原料生产策略(图1b)。对Vero细胞中P7培养基的生长动力学分析显示,培养基病毒能够有效复制,在感染后48-72小时(hpi)达到最高滴度超过7.0 log10 CCID50,感染多重度(MOI)为0.01-0.3(图1c)。为了灭活病毒产生,研究者在2-8℃,以1:40的比例将丙酸内酯与收获的病毒溶液充分混合。病毒的3批灭活消除了病毒的感染性,验证了灭活过程良好的稳定性和重复性(图1d)。Western-blotting分析显示,疫苗库中含有病毒结构蛋白(保护性抗原)(图1e)。负染色的电子显微镜图像显示椭圆形病毒颗粒,直径约为100 nm(图1f)。
BBIBP-CorV的免疫原性
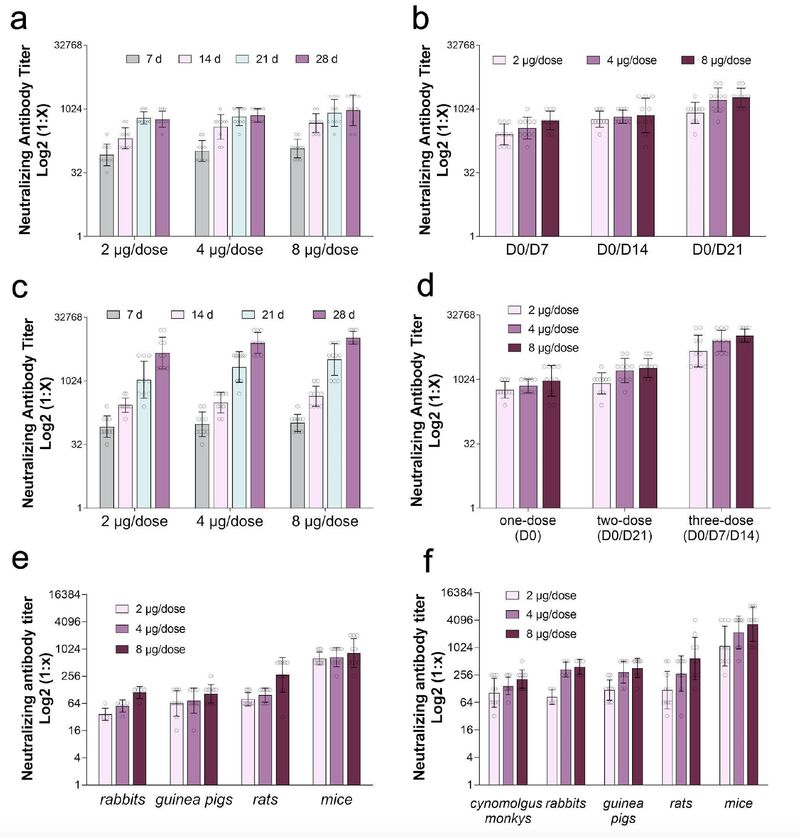
为评估BBIBP-CorV的免疫原性,研究者采用不同免疫方案和不同剂量(2、4、8μg/剂)疫苗注射BALB/c小鼠。
单剂量免疫组在第0天(D0)以三种剂量腹腔给药,分别是高(8 μg/剂)、中(4μg/剂)、低剂量(2μg/剂)的BBIBP-CorV,观察注射后7、14、21、28天中和抗体(NAb)水平。结果表明,高、中、低剂量组的血清转化速率在免疫后7天都达到100%,同时免疫效果与时间相关(图2,补充表1) 。低、中剂量组的中和抗体水平在7、14和21天显示显著的变化,但21天到28天之间无显著差异。在高剂量组中,仅在7 、14天观察到显著变化(图2a)。
双剂免疫组采用不同免疫方案(第0天/第7天、第0天/第14天、第0天/第21天分别进行免疫)。双剂量免疫中的高、中、低剂量组在第二次免疫后7天的血清阳性达到100%(图2 b,补充表1)。高、中剂量的双剂免疫方案的免疫原性明显高于单剂量免疫方案。采用第0天/第21天方案的第二次免疫后7天,其中和抗体水平最高。
研究者还测试了三剂免疫程序的免疫原性。他们在0天、7天和时14 天在小鼠腹腔接种三种剂量的疫苗,分别是高(8μg /剂)、中(4μg /剂)或低(2μg /剂)疫苗 (图2c)。在第7、14、21、28天测定各组的中和抗体水平,在第一次免疫后第7天,三组血清转化率均达到100%(图2c,补充表1)。
结果显示,三剂 (0天/7天/14天)免疫接种方案在第28天时的中和抗体水平,均高一剂免疫接种方案(图2a和2c)。此外,他们分析了单剂、双剂(0天/21天)和3剂(0天/7天/ 14天)免疫接种小鼠采用高、中、低剂量疫苗的中和抗体水平,并在第一次免疫后的28天检测中和抗体水平,以保持相同的起止点。结果显示,三剂量(0天/7天/14天)免疫接种方案的免疫原性高于单剂量和双剂量免疫接种方案(图2d)。
接下来他们在兔子、豚鼠、大鼠和小鼠等不同的动物模型中测量BBIBP-CorV的免疫原性。采用单剂量(D0)免疫接种方案对动物进行高剂量(8μg/剂)、中剂量(4μg/剂))和低剂量(2μg/剂))疫苗免疫,免疫后21天测定中和抗体水平。结果表明BBIBP-CorV具有良好的免疫原性,免疫后21天在所有动物模型中血清转化率达到100%,(图2 e,补充表1)。在三剂(0天 / 7天 / 14天)免疫组,食蟹猴、兔子、豚鼠、大鼠和小鼠接种高(8μg /剂量)中(4μg /剂量)或低(2μg /剂量)剂量的疫苗。免疫后21天在所有动物模型中血清转化率达到100%。第一次免疫后21天的中和抗体水平表明,在兔和豚鼠模型中三剂(0天/7天/14天)免疫程序的高、中、低剂量下,均高于单剂(0天)免疫程序 (图2 e 2 f,补充表1)。
非人灵长类动物模型中的保护作用
研究团队评估了BBIBP-CorV对恒河猴的免疫原性和保护作用。
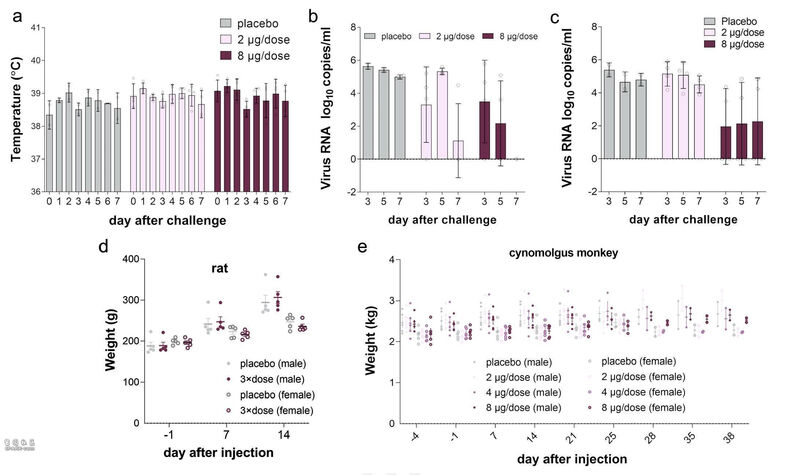
所有的恒河猴在第0天(D0)和第14天(D14)免疫2次。安慰剂组肌肉注射生理盐水,两组实验组肌肉注射低剂量(2μg/剂)或高剂量(8μg/剂)BBIBP-CorV(图3a)。病毒攻毒前,低剂量组和高剂量组的NAb GMT分别达到215和256(图3b)。在D24(第二次免疫后第10天),所有恒河猴在麻醉下经气管注射l06 TCID50的SARS-CoV-2。病毒感染后0 至7天,接种组和安慰剂组的体温在范围内波动(图3c,补充图4a)。
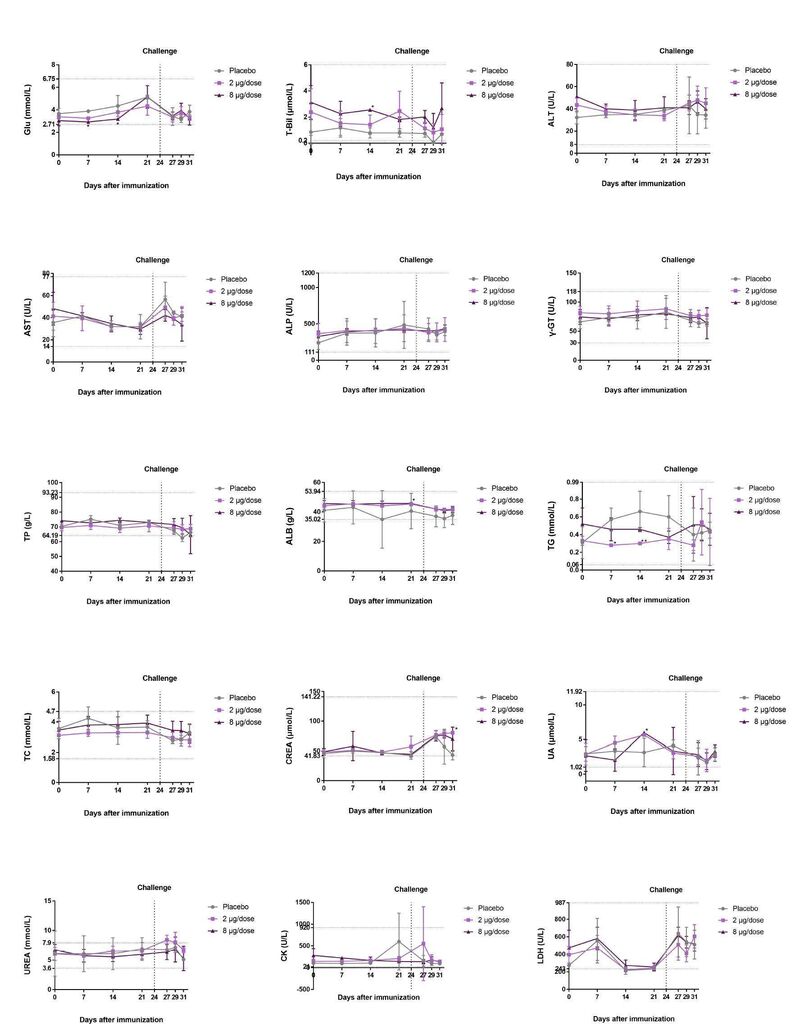
另外,恒河猴在攻毒后血清生化参数保持不变(补充图3),这一结果表明,接种BBIBP-CorV不会在血清生化参数方面显示出副作用。
接下来研究团队用RT-PCR测定了恒河猴咽喉和肛门拭子的病毒载量。结果显示,所有安慰剂组恒河猴在攻毒后整个评估期间均显示并维持高病毒载量(图3d、3e,补充图4b和4c)。
低剂量组恒河猴咽喉拭子病毒载量则在5dpi时达到峰值(5.33 log10拷贝/毫升),7 dpi时下降至1.12 log10拷贝/毫升,明显低于安慰剂组。值得注意的是,低剂量组的4只恒河猴中有3只在7 dpi时已无法检测病毒载量。而高剂量组4只恒河猴咽喉拭子病毒载量均为阴性。另外,高剂量组4只恒河猴中的2只肛门拭子也未检测到病毒载量。
7 dpi时,研究团队对所有动物实施了安乐死,以确定肺组织中的病毒载量并进行病理检查(图3f、3g)。在低剂量和高剂量组中,所有恒河猴在任何肺叶中均未检测到病毒载量,这与安慰剂组的结果有显著差异(图3f)。安慰剂组在左肺、右肺和右副肺检测到高病毒载量,病理组织学分析为严重间质性肺炎。
然而值得注意的是,在安慰剂组的7个肺叶切片中,只有3个被检测出有感染。论文指出,这可能是因为病毒在肺叶中的感染是动态变化的。
总的来说,接种BBIBP-CorV疫苗后,所有恒河猴的肺均正常,少数肺叶有局部轻度组织病理学改变(图3g),这说明接种BBIBP-CorV疫苗可有效阻断猴子SARS-CoV-2感染。
7dpi时,接受安慰剂治疗的恒河猴产生低水平的NAb,滴度为1:16,而高剂量组NAb水平最高可达1:2048(平均1:860),低剂量组NAb水平最高达1:1024(平均1:512)(图3b)。研究指出,这些结果表明,低剂量和高剂量的BBIBP-CorV对恒河猴SARS-CoV-2具有高效的保护作用,且没有观察到抗体依赖性增强感染。
安全性
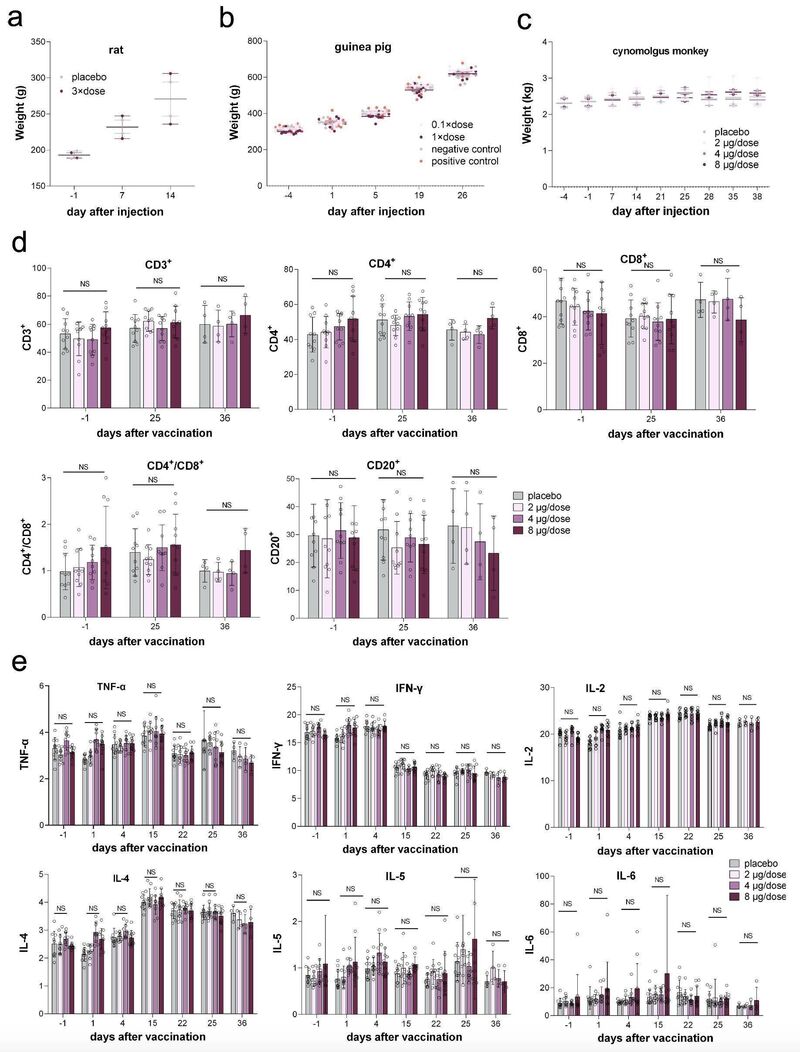
研究团队首先对大鼠(Sprague-Dawley)进行了单次肌肉注射实验,以评估BBIBP-CorV的急性毒性。研究将20只大鼠分为2组(n=10, 5/性别),肌肉注射3倍剂量BBIBP-CorV (8μg/剂,24μg/大鼠),生理盐水作为对照组。接种后连续观察14天,第15天实施安乐死,评估系统解剖和观察。
四组大鼠接种后连续14天内未见死亡或即将死亡,也未见明显临床症状。此外,实验组与对照组在体重和喂养状态上均无显著差异(图4a,补充图4d)。安乐死后这些大鼠无组织病理学改变。
值得注意的是,大鼠单次肌肉注射的最大耐受剂量(MTD)为24μg/大鼠,相当于人体的900倍。研究团队认为,这表明BBIBP-CorV对人体有潜在的良好安全性。
研究团队随后通过豚鼠肌肉注射和静脉注射评估BBIBP-CorV引起的全身过敏反应。通过临床观察和测量豚鼠体重,结果显示敏化期间未见异常反应(图4b)。阴性对照组(生理盐水)和实验组在D19、D26均未发现过敏反应症状。阳性对照组(人血白蛋白)过敏反应高度阳性(1/6只动物为阳性,3/6只动物为强阳性,2/6只动物为极强阳性)。与之形成鲜明对比的是,低、高剂量试验组D19、D26均无过敏反应,且过敏反应均为阴性。
研究团队进一步评价了BBIBP-CorV对恒河猴的长期毒性。40只猕猴(20/性别)被分成4组(5/性别/组),分别肌内注射生理盐水(组1)或2μg、4μg、8μg BBIBP-CorV(组2-4)。每组分别有3只在D25解剖,其余2只在D36解剖,进行系统解剖和组织病理检查。
在试验期间,组2-4没有出现死亡或即将死亡,淋巴细胞亚群分布(CD3+,CD3+CD4+,CD3+CD8+,CD20+,CD3+CD4+/CD3+CD8+)、细胞因子(TNF-αIFN-γ,IL-2,IL-4,IL-5和IL-6)、c反应蛋白、或体重观察也没有重大异常临床生理和病理指标 (图4c-4e,补充图4e)。
D25和D36时各剂量组的安乐死猕猴系统解剖未见异常。D25时组2-4均出现了肉芽肿性炎症,并在恢复期(D36)结束时依然存在,但与D25相比略有改善。猕猴只表现出局部刺激,表现为轻度到重度肉芽肿性炎症,但在注射后两周没有这种反应。8μg/剂实验组表现出未观察到的不良反应水平(NOAEL)。